ОСОБЕННОСТИ ОПЫЛЕНИЯ И ОПЛОДОТВОРЕНИЯ У ЯБЛОНИ В УСЛОВИЯХ ЗАКАРПАТЬЯ.
Й.М.Чернеки
Большинство закарпатских сортов яблони при самоопылении — бесплодны. Для нормального оплодотворения и получения полноценных плодов нужен перенос пыльцы с цветка одного сорта на цветки другого.
И. В. Мичурин (1948) отмечал, что перекрестное опыление у яблони ведет к резкому повышению количества плодов и улучшению их качества. Перекрестно опыляемые сорта яблонь плодоносят в том случае, если посадки их проведены совместно с сортами — опылителями.
Помимо биологических особенностей растений на результаты опыления в значительной степени влияют и погодные условия, в основном температура и влажность воздуха. В холодную дождливую погоду процент опыляемых цветков незначительный. Теплая, сопровождающаяся выпадением ночной росы погода в период цветения благоприятствует нормальному опылению. Пестики в этом случае не подсыхают и дольше сохраняют восприимчивость для принятия пыльцы.
Губительны в период цветения заморозки и дожди со снегом. Такая погода в Закарпатье зафиксирована нами 21 апреля 1962 г. Проведенные наблюдения показали, что пестики цветков, направленные вверх, более чем на 80% погибли (рис.1). Сохранились только те завязи и цветки, которые открылись несколькими днями позже или размещались с нижней стороны веток.
Пыльцевые зерна в естественных условиях с задних лапок пчелы попадают в покрывающую рыльца жидкость. Наблюдение и исследование снятой с лапок пчел пыльцы под микроскопом показало, что она очень неоднородна: встречаются крупные и довольно мелкие, разной формы пыльцевые зерна. Наблюдения позволяют предположить, что в естественных условиях может происходить опыление пчелиными не только пыльцой различных сортов яблони, а также возможно и чужеродное опыление.
Пыльца, попавшая на рыльца пестика, через 3— 4 часа начинает прорастать. Как отмечает Ф. Кобель (1957), уже через несколько часов из ростковой поры пыльцевого зерна на рыльце пестика яблони вырастает пыльцевая трубка, кончик которой наполнен густой протоплазмой и содержит вегетативное ядро и генеративную клетку.
На отдельных рыльцах таких сортов, как Батул и Джонатан, нами насчитывалось в естественных условиях более чем 30 проросших пыльцевых зерен (рис. 2а). На микротомных срезах пестиков рядом с проросшими пыльцевыми зернами отмечали и просто трехгранную пыльцу, не имеющую живого содержимого (рис. 2б). Можно предполагать, что насекомыми переносится вместе с фертильной и мертвая, нежизненная пыльца. Подсчеты показали, что из 35 пыльцевых зерен не проросло всего 3—4. Как правило, они не образовали пыльцевых трубок и были пустыми.
Пыльцевые трубки на рыльцах проросших пыльцевых зерен, благодаря положительному хемотропизму, врастают в проводниковую ткань столбика (рис. 2в).
В отличие от культуры на искусственной питательной среде, рост пыльцевой трубки в тканях пестика стимулируют и ферменты, с помощью которых пыльцевая трубка для своего роста и развития растворяет находящиеся в проводниковой ткани запасные вещества. Исследования Петона (Paton I. В., 1921) на сибирской яблони показали, что в пыльцевых трубках дикой яблони имеются ферменты амилазы, каталазы, редуктазы, пектинтазы, зимазы и инвертина.

Рис. 2. а — Рыльце пестика, усеянное пыльцевыми зернами; б — стерильная пыльца на рыльце; в — прорастание пыльцевых зёрен.
Пыльцевые трубки в семенных камерах растут вдоль стенок, из клеток которых они, очевидно, способны извлекать питательные вещества и вызывают, по наблюдениям П. Штейнеггера (Steinegger Р., 1922), разбухание отмеченных выше клеток.
В связи с анатропным размещением семяпочек у яблони пыльцевая трубка быстро достигает микропиле. Таким образом, она приводит оплодотворяющие элементы к зародышевому мешку.
В обычных условиях, отмечает Ф. Кобель (1957), в микропиле проникает лишь одна пыльцевая трубка. Нам удалось наблюдать присутствие нескольких пыльцевых трубок в микропилярной части семяпочки.
Следует отметить, что некоторые исследователи (А. А. Чеботарь, 1960) наблюдали пыльцевые трубки в столбике пестика, которые сливались в общий «поток или русло». Такой поток трубок обладает большими энергетическими возможностями, чем одиночные.
На препаратах таких сортов, как Астраханская красная, Батул и Джонатан, мы часто отмечали 8—10 пыльцевых трубок, они образовывали в пестике целую систему каналов (рис. 3а). Большое количество трубок гарантирует проведение оплодотворяющих элементов до зародышевого мешка, что обусловливает и множественные оплодотворения. Поэтому в естественных условиях не исключена возможность проникновения одновременно двух пыльцевых трубок в один зародышевый мешок, нередко разных сортов, и яйцеклетка может оплодотвориться спермием из одной пыльцевой трубки, а полярные ядра — спермием из другой.
А. Остервальдер (1910) наблюдал процесс оплодотворения зародышевого мешка яблони примерно через 4 дня, а П. Штейнеггер (1922) в своей работе отмечал, что пыльцевые трубки достигали зародышевого мешка через 4—6 дней после опыления. Однако они не указывают на особенности отдельных сортов и на влияние скорости роста пыльцевых трубок при само- или перекрестном опылении.
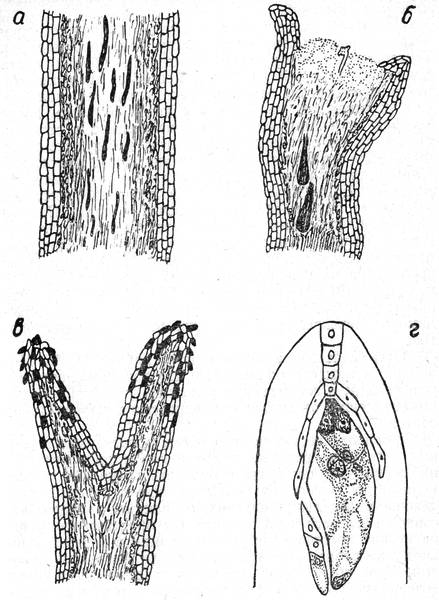
Рис. 3. а — система каналов пыльцевых трубок в столбике пестика; б — образование вздутий на кончиках пыльцевых трубок при самоопылении у сорта Батул; в — мертвые клетки на рыльцах пестиков; г — двойное оплодотворение элементов зародышевого мешка у яблони.
Наши наблюдения показали, что эти процессы протекают у разных сортов по-разному. Так, например, при искусственном перекрестном опылении яблони сорта Джонатан пыльцой Делишеса красного мы отмечали, что оплодотворение зародышевого мешка наступало через 48 часов, но не более чем через 56 часов после их искусственного опыления. Значительно отставали, по сравнению с предыдущим опытом, в быстроте роста пыльцевые трубки при самоопылении таких сортов, как Пармен зимний золотой, триплодный сорт Ренет канадский. Даже через 6—8 дней после самоопыления пыльцевые трубки вблизи семяпочек у сорта Пармен зимний золотой нами не наблюдались, хотя за это время уже большинство зародышевых мешков обнаружили признаки дегенерации. Совершенно не росли пыльцевые трубки (на их кончиках появились вздутия) в столбиках при самоопылении у сорта Батул (рис. 3б). Эти опыты нам эмбриологически подтверждают о необходимости перекрестного опыления вышеуказанных сортов.
Здесь целесообразно вспомнить работу И. В. Мичурина, в которой указывается, что при опылении смесью пыльцы в естественных условиях элементы зародышевого мешка отдают, свое предпочтение пыльце диких сортов.
Эмбриологические исследования, проведенные в горных условиях, свидетельствуют о целом ряде отклонений в процессе опыления и оплодотворения. Сравнивая количество нормально оплодотворенных зародышевых мешков в отдельных завязях, собранных в предгории, горной местности и на равнине, мы обратили внимание на низкий процент завязавшихся семян в горных условиях по сравнению с предгорьем или низинным районом. Это объясняется неблагоприятными погодными условиями, пониженной температурой, высокой влажностью и холодными дождями в период цветения, которые пагубно подействовали на рыльца пестиков, отчего те дегенерировали и содержали многочисленные мертвые клетки (рис. 3в).
В благоприятных условиях пыльцевая трубка, как правило, внедряется в зародышевый мешок в ее микропилярной части. Как отмечает А. В. Константинов (1958), часто пыльцевая трубка проходит через одну из синергид, разрушая ее. Ф. Кобель считает, что в момент оплодотворения синергиды претерпевают распад. В действительности же в оплодотворенных зародышевых мешках нам не удалось наблюдать нормальные синергиды, так как они находились в это время в стадии дегенерации (рис. Зг).
Оплодотворение зародышевого мешка у яблони происходит, в основном, как описано С. Г. Навашиным (1898) и дополнено С.Я.Модилевским (1953) для покрытосеменных растений: содержимое пыльцевой трубки изливается несколько ниже яйцеклетки, ближе к центральной клетке зародышевого мешка, содержащей полярные ядра. Один из спермиев направляется к яйцеклетке и сливается с ней. Второй — вступает в контакт с полярными ядрами, при этом его ядро сливается с ними, а цитоплазма входит в состав центральной клетки зародышевого мешка. Зародышевый мешок одновременно всасывает и содержимое пыльцевой трубки, как и протопласты синергид, и обычно остатки антипод.
Нельзя согласиться с высказыванием А. Остервальдера и Штейнеггера или даже Ф. Кобеля о том, что оплодотворение сводится к простому суммированию половых клеток, или, как А. Остервальдер считал, слиянию одних лишь половых ядер.
Многие советские исследователи (Я. Е. Элленгорн, 1949; В. В. Светозарова, 1950; А. С. Афанасьева, 1956) доказали, что объяснять двойное оплодотворение простым слиянием половых клеток никак нельзя. Имеется целый ряд данных, свидетельствующих о влиянии плазмы и оплодотворяющих элементов на семена и плоды.
Процесс оплодотворения у яблони, как и во всех цветковых растениях, является биологически сложным, при котором происходят биохимические процессы ассимиляции и усвоения веществ, представляющих к половому процессу родительские пары. Как отмечает Е. Н. Герасимова-Навашина (1942), даже так называемая «вегетативная плазма» пыльцевой трубки, которой приписывалось до сих пор самое большее — питательные функции, должна быть признана источником живого вещества, способствующего развитию зародыша и эндосперма, специфически изменяющего ход этого развития.
В заключение следует отметить, что оплодотворенная яйцеклетка после ассимиляции содержимого пыльцевой трубки становится крупнее, а протоплазма — более плотной. Неоплодотворенная, наоборот, через 5—6 дней после цветения начинает дегенерировать. Если это разрушение не повлияет на нуцеллус, то в таком случае, благодаря развитию новых археспориальных клеток, могут развиваться дополнительно апоспорические зародышевые мешки.
РЕЗЮМЕ
Сложность климатических условий Закарпатской области накладывает свои отпечатки на процессы опыления и оплодотворения яблони. Наблюдения и эксперименты проводились на местных и широко распространенных сортах (Батул, Делишес, Джонатан и др.)
Весенние заморозки в период цветения и понижения температур губят рыльца пестиков и отрицательно влияют на генеративные органы плодовых, в результате чего образуются стерильные пыльцевые зерна и дегенерируют элементы зародышевого мешка.
Приспосабливаясь к отрицательному влиянию климатических факторов, плодовые растения приобретают более 30 пыльцевых зерен на рыльцах. В результате развиваются дополнительные зародышевые мешки, которые обеспечивают нормальное развитие репродуктивных органов яблони.
ЛИТЕРАТУРА
Афанасьева А. С. Некоторые новые данные о процессе оплодотворения у пшеницы. Общая биология, №1, 1956.
Герасимова-Навашина Е. И. К цитолого-эмбриологическому пониманию процесса опыления. Морф. и анат. растений Изд. АН СССР, 1952.
Кобель Ф. Плодоводство на физиологической основе. Сельхозгиз, М., 1957.
Константинов А. В. Процесс оплодотворения у яблони и ранний эмбриогенез. ВИР. Л., 1958.
Константинов А. В. Эмбриология некоторых сортов яблони. Изд. АН СССР, № 2, 1960.
Мичурин И. В. Сочинения. Сельхозгиз, М., 1948.
Модилевский Я. С. Эмбриология покрытосеменных растений. Киев, 1953.
Поддубная-Арнольди В. A. Материалы к истории исследования процесса оплодотворения у покрытосеменных растений. Труды ГБС, т. VI, М., 1952.
Элленгорн Я. E., Светозарова В. В. Процесс оплодотворения покрытосеменных растений. Изд. АН СССР, № 3, 1959.
Osterwalder О. А. Blütenbiologie, Embriologie und Entwiclung der Frucht unserer Kernobstraune. Land. Jahrb. 1910.
Paton J. B. Pollen and enzymes. Amer. Bot. Jour. 8. № 10, 1920.
Steinegger Р. Cytologisch Bedingte Ei und Zygotensterilität bei triploiden Apfelsorten. Zürich. 1922.
Источник: Вопросы охраны природы Карпат. Карпаты. Ужгород, 1968
Комментировать